
Keratella tropica - Tropisch puzzelraderdier

WETENSCHAPPELIJKE NAAM
Keratella tropica (Apstein, 1907)
In het zuiden van Frankrijk en Spanje is het Tropisch puzzelraderdier inheems [2]. De soort komt wereldwijd voor in tropisch en subtropisch zoetwater [3], maar kan tijdens de zomer ook gedijen in gematigde streken. Het is een vrijzwemmende soort die ook brak water tolereert [4].
Eerste waarneming in België
De eerste waarneming van dit raderdiertje in België vond plaats in 1974, in het Donkmeer (Overmere) [5]. In datzelfde jaar werd het Tropisch puzzelraderdier ook aangetroffen in de Watersportbaan (Gent) [6].
Verspreiding in België
Wetenschappers meldden in 1983 dat het Tropisch puzzelraderdier in België een invasief karakter heeft tijdens hete zomers [7]. In de lente en zomer van 2002 werd de diversiteit van de raderdiertjes in het Schelde-estuarium specifiek onderzocht. Het Tropisch puzzelraderdier werd er toen waargenomen, telkens in juli en augustus, en veelal in de bovenloop van de Schelde tot Gent. Maar ook ter hoogte van Antwerpen – in het brak waterdeel van het estuarium – trof men enkele exemplaren van dit raderdiertje aan [4].
Verspreiding in onze buurlanden
In Nederland werd het voorkomen van dit raderdiertje voor het eerst gemeld in augustus 1959, toen het werd aangetroffen in de zoetwatergetijdenzone van de monding van de Rijn, ter hoogte van Biesbosch [8]. Later, in 1976 en 1977, werd deze soort opnieuw gesignaleerd in respectievelijk het Hollands Diep ten zuiden van Rotterdam en in de Reewijkse Plassen tussen Rotterdam en Amsterdam [6].
Dit raderdiertje komt ook voor in zoetwater in Frankrijk. In het zuiden van Frankrijk en Spanje is de soort echter inheems [2]. In Zuid-Frankrijk werd dit radardiertje gevonden in de provincies Landes en Gironde (in het zuidwesten) en in La Dombes (ten noorden van Lyon) [9]. Verder werd dit diertje tussen 1972-1975 en 1987-1988 door onderzoekers waargenomen in kunstmatige reservoirs verspreid over gans Spanje [10, 11]. Ook in natuurlijke waterlichamen wordt deze soort aangetroffen [12]. Vandaag wordt dit diertje beschouwd als een algemene zoetwatersoort binnen Europa [4].
De introductie gebeurde mogelijks via het ballastwater van schepen. Naast het internationaal transport kan ook het lokaal scheepvaartverkeer een rol hebben gespeeld in de verspreiding van deze exoot [13]. In het geval van de Schelde kan de introductie ook hebben plaatsgevonden via transport over de kanalen vanuit de Rijn of de Maas [4].
Watervogels – zoals de wilde eend – kunnen eveneens een rol spelen in de verspreiding van raderdiertjes, en dit op twee manieren. Enerzijds kunnen de rusteieren van raderdiertjes opgegeten worden en op een andere plaats – samen met de uitwerpselen – vrijkomen. Anderzijds kunnen de diertjes – of hun rusteieren – blijven kleven aan de poten of de veren van vogels en zo naar andere gebieden worden overgebracht [14]. De locaties waar het Tropisch puzzelraderdiertje gevonden werd, liggen op de migratieroutes van vogels die overwinteren in (sub)tropische gebieden en broeden in koude en gematigde streken [9].Het transport via ballastwater van schepen of de verspreiding via migrerende vogels kan zorgen voor een snelle uitbreiding van het areaal van deze soort richting verschillende zoetwaterlichamen en estuaria [4, 9].
Daarbovenop kan dit diertje zich snel op geslachtelijke en ongeslachtelijke wijze voortplanten, waardoor uit één enkel rustei een nieuwe populatie kan ontstaan [15].Zoals de meeste raderdiertjes komt het Tropisch puzzelraderdier vooral voor in zoet water. De soort kan echter ook overleven in het brakke milieu. In de Schelde is dit diertje tot een zoutgehalte van 2,62 psu geobserveerd. Op het moment van zijn aanwezigheid varieerde de temperatuur er tussen 18,5 en 23,6 °C, eerder warme condities voor de Schelde. Deze soort is in onze streken enkel in de zomermaanden in grote aantallen te vinden of op plaatsen waar het water verwarmd wordt door warm koelwater [4, 6, 8]. De opwarming van de aarde werkt dan ook de ontwikkeling van deze soorten in onze regio’s in de hand.
Daarenboven blijkt dit raderdiertje goed te gedijen in voedselrijke omstandigheden, typisch voor estuaria zoals de Schelde [16, 17].
In de zomermaanden, vooral in augustus, kan het Tropisch puzzelraderdiertje tot bijna 8% van de totale densiteit van alle raderdiertjes uitmaken in het Schelde-estuarium. Dit kan mogelijk een effect hebben op het voedselweb van het estuarium [4], hoewel daar nog geen bewijzen voor zijn.
Raderdiertjes zijn micro-organismen, enkel te herkennen met behulp van een microscoop. Ze hebben een vooraan gelegen kop, een romp en een achteraan gelegen regio of voet (figuur 1). De kop draagt een gewimperde structuur, het raderorgaan of de corona, dat gebruikt wordt om zich te voeden, door partikels uit het water te filtreren, en om zich voort te bewegen. Te midden deze structuur bevindt zich de mond en net daarachter de mastax, een harde structuur die gebruikt wordt om de voedselpartikels te kauwen. De vorm van de mastax is vrij specifiek voor iedere soort, en kan gebruikt worden als identificatiekenmerk. Sommige soorten zijn omgeven door een pantser of ‘lorica’, terwijl dit bij anderen ontbreekt [18].

Figuur 1: Algemeen bouwplan raderdieren (Bron: VLIZ).
Het Tropisch puzzelraderdier is ongeveer 165 μm groot (stekels meegerekend) (figuur 2). Het diertje is omgeven door een pantser, dat gebruikt wordt voor de identificatie op soortniveau. Het bestaat uit verschillende platen, waarvan de aanwezigheid van een klein achterwaarts gelegen plaatje de soort karakteriseert. Verder heeft het pantser zes stekels aan de kopzijde en twee stekels achteraan [4], maar de lengte ervan varieert sterk tussen individuen. Eén van deze twee stekels (de rechterstekel) is steeds langer dan de andere stekel [19] (figuur 3). Puzzelraderdiertjes hebben geen voet [2].

Figuur 2: Het Tropisch puzzelraderdier (Bron: Azémar et al. 2007) [3, 4].

Figuur 3: Variatie in grootte van de rechterstekel bij het Tropisch puzzelraderdier (Bron: Azémar et al. 2007) [4].
Onderzoek wijst uit dat een vrouwelijk raderdiertje de aanwezigheid van een predator in de omgeving kan detecteren via bepaalde stoffen (‘cairomonen’) die de predatoren afgeven. Als reactie hierop gaan de nakomelingen – bij ongeslachtelijke voortplanting via amictische eitjes (zie verder) – stekels ontwikkelen die als verdediging tegen de aanwezige predatoren dienstdoen. Wetenschappers noemen dit fenomeen ‘fenotypische plasticiteit’, wat betekent dat het vermogen om deze stekels te ontwikkelen aanwezig is in het genetisch materiaal (DNA), maar dat de eventuele ontwikkeling van deze stekels bepaald wordt door de omgeving [2, 20].
Het Tropisch puzzelraderdiertje behoort tot de groep van de Monogononta. Deze naam verwijst naar de aanwezigheid van slechts één (mono) geslachtsklier (gonade). Bij de raderdieren zijn er altijd veel meer vrouwelijke individuen dan mannelijke, waarbij de mannetjes veel kleiner zijn. Als er weinig mannetjes zijn, kunnen vrouwtjes zich zonder bevruchting voortplanten, ook wel parthenogenese of ‘maagdelijke voortplanting’ genoemd. Hierbij leggen ze eitjes die men ‘amictisch’ noemt, dit wil zeggen dat ze twee exemplaren van elke chromosoom bevatten (2N). Daarnaast leggen vrouwtjes, bij verslechterende omstandigheden, ook ‘mictische’ eitjes die slechts één stel chromosomen bevatten (N), al is de situatie hier iets complexer. Een deel van de mictisch eitjes wordt niet bevrucht en deze eitjes evolueren tot mannetjes, die vervolgens de andere mictische eitjes bevruchten [2, 15]. Uit een bevrucht mictische eitje ontwikkelt zich een ‘rustei’ of ‘duurei’. Dit ei zal pas bij betere omstandigheden – soms pas na enkele maanden tot zelfs jaren – uitkomen en zal altijd vrouwelijke individuen voortbrengen. Deze duureieren zijn heel geschikt om minder gunstige milieuomstandigheden te overbruggen en kunnen bovendien met de stromingen naar andere gebieden getransporteerd worden (figuur 4).
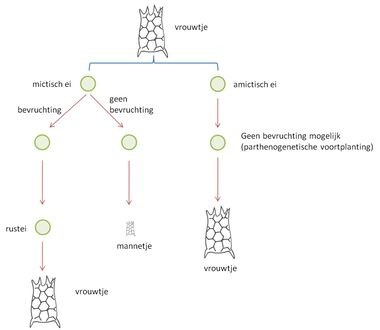
Figuur 4: Geslachtelijke en ongeslachtelijke voortplanting bij raderdieren (Bron: VLIZ).
[1] World Register of Marine Species (WoRMS) (2020). Keratella tropica (Apstein, 1907). [http://www.marinespecies.org/aphia.php?p=taxdetails&id=248010] (2020-11-17).
[2] Segers, H. (2009-2011). Persoonlijke mededeling
[3] Segers, H. (2007). Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 1564: 1-104. [http://www.vliz.be/en/imis?module=ref&refid=140748]
[4] Azémar, F.; Van Damme, S.; Meire, P.; Tackx, M. (2007). New occurrence of Lecane decipiens (Murray, 1913) and some other alien rotifers in the Schelde estuary (Belgium). Belg. J. Zool. 137(1): 75-83. [http://www.vliz.be/en/imis?module=ref&refid=112260]
[5] Coussement, M. (1977). Nieuwe gegevens omtrent de Rotatoria-fauna van het Donkmeer in Oost-Vlaanderen. Natuurwet. Tijdschr. 58(3): 138-146. [http://www.vliz.be/en/imis?module=ref&refid=140153]
[6] Leentvaar, P. (1980). Note on some Brachionidae (Rotifers) from the Netherlands. Hydrobiologia 73(1-3): 259-262. [http://www.vliz.be/en/imis?module=ref&refid=139914]
[7] Dumont, H.J. (1983). Biogeography of rotifers. Hydrobiologia 104: 19-30. [http://www.vliz.be/en/imis?module=ref&refid=139918]
[8] Leentvaar, P. (1961). Quelques rotateurs rares observes en Hollande. Hydrobiologia 18: 245-251. [http://www.vliz.be/en/imis?module=ref&refid=140138]
[9] De Ridder, M. (1981). Some considerations on the geographical distribution of rotifers. Hydrobiologia 85: 209-225. [http://www.vliz.be/en/imis?module=ref&refid=139912]
[10] Guiset, A. (1977). General distribution of planktonic rotifers in Spanish reservoirs. Ergeb. Limnol. 8: 222-225. [http://www.vliz.be/en/imis?module=ref&refid=140334]
[11] De Manuel Barrabin, J. (2000). The rotifers of Spanish reservoirs: Ecological, Systematical and zoogeographical remarks. Limnetica 19: 91-167. [http://www.vliz.be/en/imis?module=ref&refid=140744]
[12] Miracle, M.R. (1982). Biogeography of the freshwater zooplanktoninc communities of Spain. J. Biogeogr. 9(6): 455-467. [http://www.vliz.be/en/imis?module=ref&refid=142407]
[13] Wasson, K.; Zabin, C.J.; Bedinger, L.; Diaz, M.C.; Pearse, J.S. (2001). Biological invasions of estuaries without international shipping: the importance of intraregional transport. Biol. Conserv. 102: 143-153. [http://www.vliz.be/en/imis?module=ref&refid=140750]
[14] Frisch, D.; Green, A.J.; Figuerola, J. (2007). High dispersal capacity of a broad spectrum of aquatic invertebrates via waterbirds. Aquat. Sci. 69: 568-574. [http://www.vliz.be/en/imis?module=ref&refid=140746]
[15] Thane, A. (1974). Rotifera, in: Giese, A.C. et al. Reproduction of marine invertebrates: 1. Acoelomate and Pseudocoelomate Metazoans. Reproduction of marine invertebrates, 1. Academic Press: New York: pp. 471-484. [http://www.vliz.be/en/imis?module=ref&refid=10016]
[16] Duggan, I.C.; Green, J.D.; Shiel, R.J. (2002). Distribution of rotifer assemblages in North Island, New Zealand, lakes: relationships to environmental and historical factors. Freshwat. Biol. 47: 195-206. [http://www.vliz.be/en/imis?module=ref&refid=140126]
[17] Van Damme, S.; Struyf, E.; Maris, T.; Ysebaert, T.; Dehairs, F.; Tackx, M.; Heip, C.; Meire, P. (2005). Spatial and temporal patterns of water quality along the estuarine salinity gradient of the Scheldt estuary (Belgium and The Netherlands): results of an integrated monitoring approach. Hydrobiologia 540(1-3): 29-45. [http://www.vliz.be/en/imis?module=ref&refid=75934]
[18] Ruppert, E.E.; Barnes, R.D. (1994). Invertebrate zoology. 6th edition. Saunders College Publishing: Orlando. ISBN 0-03-026668-8. 1056 pp. [http://www.vliz.be/nl/catalogus?module=ref&refid=9414]
[19] Green, J. (1980). Asymmetry and variation in Keratella tropica. Hydrobiologia 73(1-3): 241-248. [http://www.vliz.be/en/imis?module=ref&refid=139916]
[20] Zagarese, H.E.; Marinone, M.C. (1992). Induction and inhibition of spine development in the rotifer Keratella tropica. Freshwat. Biol. 28: 289-300. [http://www.vliz.be/en/imis?module=ref&refid=140341]