
| Trophos |
| Higher trophic levels in the Southern North Sea |
|
||||
| |
|
|
|
| home | objectives | results | background | activities | databases | partners |
The Belgian Continental Shelf (BCS) is characterized by numerous subtidal sandbanks. Their close position to the coastline and the shallowness of the area make them unique, both from an ecological and geological point of view. It has recently been shown that the sandbank system in the Belgian coastal area acts as an internationally important resting and foraging area for various seabird species.
The TROPHOS project was carried out in the framework of the SPSD-II research action and is therefore closely linked to other SPSD-II projects like AMORE-II, BALANS and MAREBASSE. TROPHOS also runs in close interaction with other research projects that are being carried out at the Marine Biology Section in Ghent. Among them are the HABITAT project, the IWT projects 'Spatio-temporal variability and population dynamics of the Abra alba - Mysella bidentata community on the Belgian Continental Shelf.' and 'Marine Biodiversity: a molecular approach' and the FWO-project on 'Dispersion of Clupeidae'.
The different research topics that were studied in Trophos project are:The former SPSD-I research project 'Structural and functional biodiversity of North Sea ecosystems: species and their habitats as indicators for a sustainable development of the 'Belgian Continental Shelf' (1997-2001) aimed to assess the factors that determine and influence the marine biodiversity of the Belgian Continental Shelf.
The different research topics that were studied in this former project are:
Within that project, an emphasis was made to incorporate all existing and new biodiversity information (community, population and genetic level) on all benthic compartments, including demersal fish, their parasites and birds. Most emphasis was given on the description of the structural aspects of benthic biodiversity in order to find a pattern within the relationship between the biological and the geochemical characteristics of the sediment. The question whether the structuring of ecosystem and species diversity and its functioning is reflected in the genetic structuring of marine taxa on the Belgian Continental Shelf was addressed for the first time.
- Monitoring of the Belgian Continental Shelf
- Spatial patterns on sandbank ecosystems
- Ecological indicators
- Ecological sensitive areas
- Bentho-pelagic coupling
- Genetic biodiversity of ecologically important fishes
At the bottom of this page you can find a list of publications related to these research topics.
Monitoring of the Belgian Continental Shelf
Clear patterns in average species number have been observed for the meiobenthos, the macrobenthos, and the hyperbenthos. The species number of benthic assemblages in inshore waters changes dramatically between the eastern and western end of the Belgian coast.
A second gradient runs perpendicular to the coastline and involves an increasing species number and diversity offshore. A third gradient relating to depth within a zone needs yet to be confirmed.
An offshore decrease in density has also been observed for macrobenthos and hyperbenthos. Avifauna data do not confirm these patterns even though seabird assemblages offshore differ from the inshore avifauna. The monitoring of the hyperbenthos shows that the estuarine outflow on the one hand and the hydrodynamic setting and protective function of the sandbanks on the other hand create a diverse environment along the Belgian coastal zone. The sandbanks system extends moreover in gently sloping sandy beaches. This makes the setting of the Belgian coastal sandbank system ecologically unique.
Spatial patterns on sandbank ecosystems
Benthos

The meiobenthos of the Flemish Banks, the Hinder Banks and the Zeeland Banks is divided over four different communities, which do not reflect the geographical position but are influenced by local differences in sedimentological characteristics within sandbanks. The sedimentological differences influenced the taxon diversity as well. Sediments with a median grain size below 300 µm were poor in terms of densities and number of taxa, while coarser sediments were richer in taxon diversity. Sediments with a median grain size between 300-450 µm were rich both in terms of diversity and density, while sediments with a median grain size of >450 µm still showed high diversity but lower densities.
The information on the species diversity of the most dominant group within the meiobenthos, the free-living marine nematodes indicates that sandbanks can be seen as geographically isolated "islands" since tidal current patterns around these sandbanks prevent nematodes from being transported from one sandbank to another. A detailed study of the Kwintebank, which is heavily exploited for sand and gravel extraction, shows that a depression occurred in the middle of the sandbank due to gravel extraction (comparison between 1978 and 1997).
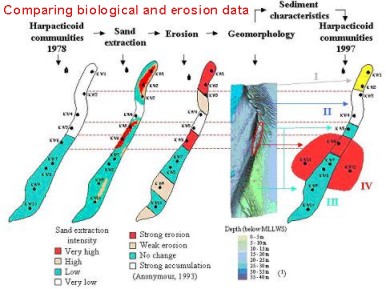
In the centre of the sandbank, copepod diversity decreased and a shift was recorded from a species rich northern community to a less divers southern community (less species, high dominance of Paraleptastacus espinulatus, Leptastacus laticaudatus s. str. and Kliopsyllus constrictus s. str.) as a result of changes in sediment characteristics. The altered sediment composition may be the result of an accretion of fine sediments, as a consequence of changed current patterns in the depression. The extension of the present depression due to sand extraction can become quite problematic if the human-induced physical disturbances may cause a continuing erosion and impoverishment.
A thorough analysis of all available data of macrobenthos of the BCS (with an extensive survey on the sandbank systems) shows that five different macrobenthic communities (1. Abra alba-Mysella bidentata; 2. Nephtys cirrosa; 3. Ophelia limacina-Glycera lapidum; 4. Eurydice pulchra-Scolelepis squamata and 5. Barnea candida community) can be distinguished with five transitional groups in between them.
All these groups are clearly linked to sediment characteristics and to their occurrence related to the geographical position on the sandbanks. These communities are ecological important, because they are a representation of the different habitats that are characteristic for the BCS.
Obviously, different macrobenthic communities and species associations show different temporal dynamics. A successful recruitment and survival of one species, for instance, will only take place in the community where this species belongs to and not in any other community. Instead of putting a lot of effort, time and money in working out time-series of lots of macrobenthic samples, randomly spread over the area under consideration, monitoring of the macrobenthos should concentrate on the spatial distribution of the macrobenthic 'potentials'. This can be done by the identification of (1) the spatial distribution of the macrobenthic communities actually present, (2) the communities' environmental 'needs', and, finally, (3) the communities' 'potentials' by means of long-term monitoring of some selected stations within each community.
Parasites
The study concentrates on ectoparasites belonging to the genus Gyrodactylus (Monogenea, Platyhelminthes) parasitizing the threespine stickleback (Gasterosteus aculeatus) and gobies from the genus Pomatoschistus (Gobioidea, Teleostei). Many new species have been found. Thanks to the available molecular techniques the detection of putative new species has become considerably easier. Both morphometric and genetic analyses are used in the species descriptions.
Avifauna

Out of the 121 bird species encountered at sea during the period 1992-98, 23 were selected as being true marine species that occur in relatively high densities within the Belgian waters. Of these 23 species, six can be classified as 'focal species' (little gull, diver spp., common scoter, sandwich tern, common tern, little tern) and another five as 'locally important species'.
A cumulative picture of the abundance and conservation value of six focal species indicates that the western Coastal Banks (Trapegeer, Nieuwpoort Bank, Stroombank, Balandbank, and Smal Bank), the Vlakte van de Raan and most of the Flemish Banks are avian hotspots throughout the year. The eastern Coastal Banks (Wenduinebank and to a lesser extent the Paardenmarkt) are very important from spring till autumn, mainly as feeding grounds for the three tern species. In winter this area is of marginal importance. On comparison the deep-water zone, the Zeeland ridges and the offshore Hinder Banks are less important.
Ecological indicators
In general, characteristics (density, biomass, biodiversity, species composition) of biological communities can be linked to habitat disturbance. However, since biological communities are quite complex to investigate, a set of ecological indicators is put forward as easy measures for community change due to general disturbances. Four 'easy measures' will be discussed: 1. Nematode Biomass Spectra, 2. Occurrence of Lanice conchilega,
3. Speciation patterns in parasites and 4. Distribution patterns of seabirds.
1. Nematodes : the biomass distribution over size requires no taxonomical knowledge and can be used by non-specialist. Nematode biomass spectra (NBS) were constructed for undisturbed sandbanks, physically disturbed sandbanks (sand extraction on the Kwinte Bank), eutrophicated and less eutrophicated coastal stations and during a pulsed supply of fresh phytodetritus following a spring phytoplankton bloom. Using NBS allows in tracking changes in all the communities mentioned above.
2. Lanice conchilega : Lanice conchilega is a tube-building, suspension or deposit feeding polychaete, of which the tube has a length of up to 45 cm. Lanice conchilega typically occurs within the macrobenthos-rich Abra alba - Mysella bidentata community. On the western Coastal Banks the polychaete was also found in the Magelona mirabilis species association. Within both communities, a positive correlation between the macrobenthic density and the number of species per sample (N0), on the one hand, and the density of L. conchilega on the other hand was found. The increase in habitat complexity in dense patches of L. conchilega might be responsible for the increasing macrobenthic density and diversity.
3. Parasites: Parasite speciation is influenced by ecological and phylogenetic factors. By comparing host and parasite phylogenies a differentiation can be made between the different speciation modes.
And in the case of strict co-speciation parasite phylogeny mirrors host phylogeny. This mirror association is absent in the present study, suggesting the important influence of ecological factors in this host-parasite system.
4. Seabirds: seabirds are often used as indicators of changes in the marine environment. Occupying high trophic levels they accumulate persistent chemicals and are extremely vulnerable to their effects, making them of value as sensitive indicators of marine pollution with chemicals. Monitoring of their numbers and distribution at the BCS can be a strong tool and an early warning system for changes within the marine ecosystem. Monitoring of contaminants in tern eggs and feathers can be used as an indicator of pollution in the Belgian waters. At present it is too early to use seabirds as indicators of changes in marine prey stock (pelagic fish, bivalves) since detailed knowledge on their feeding ecology and on food-chain interactions is still missing.
Ecological sensitive areas
The Belgian sandbank system harbour important numbers of fish and macrocrustaceans such as sole (Solea solea) and the common shrimp (Crangon crangon). Young macrocrustaceans have their maximum abundance during summer. Recruitment of fish larvae is well defined in time and happens for most species during April May. The Flemish Banks have the most abundant patterns in density for the young decapods larvae. This distribution pattern can be explained on the basis of the hydrodynamical regime in the area. Older decapod larvae reach highest densities on the Coastal Banks. Also the sheltered and food-enriched Coastal Banks provides suitable adult habitats. For example, zoea larvae of common shrimp Crangon crangon reach highest densities at the Flemish Banks, its postlarval stage is most abundant at the Coastal Banks, while the adults are dominant users of the adjacent beaches. It has also been shown that postlarval stages of Crangon crangon enter the Westerschelde estuary and use the nearby tidal marshes as a nursery.
The impact of sand and gravel extraction on benthos biodiversity has been demonstrated for the occurrence of copepods on the Kwintebank. Comparison between 1978 and 1997 was made and drastic changes have been found, both in sediment characteristics and the associated composition of the copepod communities.

Oil sensitive areas for seabirds: Winter and to a lesser extent early spring turn out to be the most delicate periods for oil-pollution. And the closer an oil-slick approaches the coastline the worse the presumed impact on seabirds will be. Within each season the western sandbank systems western Coastal Banks and Flemish Banks are highly oil-sensitive. During winter the eastern Coastal Banks (with the exception of the Vlakte van de Raan) are less vulnerable than the western part of Belgian inshore waters. In spring and summer the eastern Coastal Banks are most vulnerable.
Disturbance-sensitive seabirds such as scoters, divers, auks and grebes are most abundant in Belgian marine waters in winter and early spring. Higher disturbance-sensitivity values in autumn compared to summer are due to increasing numbers of sensitive species from October onwards. Most vulnerable sites in terms of disturbance are the western Coastal Banks and Flemish Banks, both in winter and early spring.
List of relevant publications
- Al-Hassan, A. J., Webb, C. J., Giama, M. & Miller P. J. (1987). Phosphoglucose isomerase polymorphism in the common goby, Pomatoschistus microps (Kroeyer) (Teleostei: Gobiidae), around the British Isles. J. Fish Biol. 30: 281-298.
- Anonymus (1993). Effecten op het marien leefmilieu van de zand- en grindwinningen op het Belgisch Continentaal Plat. Annalen der mijnen van België, syntheseverslag, 2: 49 pp.
- Barrett, R.T. 2001. The breeding demography and egg size of North Norwegian Atlantic puffins Fratercula arctica and razorbills Alca torda during 20 years of climatic variablility. Atlantic Seabirds 3: 97-112.
- Becker, P.H., Munoz Cifuentes, J., Behrends , B., Schmieder, K.R. (2001). Contaminants in bird eggs in the Wadden Sea. Spatial and temporal trends 1991-2001. Wadden Sea Ecosystem No. 11. Common Wadden Sea Secretariat, Trilateral Monitoring and Assessment Group, Wilhelmshaven, 68 p.
- Belkhir, K. (1999). GENETIX, logiciel sous WindowsTM pour la génétique des populations. Laboratoire Génome et Populations, Université de Montpellier II.
- Beyst, B., Hostens, K., Mees, J. (2001). Factors influencing fish and macrocrustacean communities in the surf zone of sandy beaches in Belgium: temporal variation. J. Sea Res. 44: 281-294
- Boesch, D.F., Turner, R.E. (1984) Dependence of fishery species on salt marshes: the role of food and refuge. Estuaries 7: 460-468.
- Brooks D.R. & McLennan D.A. (1993). Phylogeny, ecology and behavior. The University of Chicago Press, Chicago.
- Bulteel, G., van der Vloet, H. (1969). Zeevogelwaarnemingen aan de Belgische kust. Giervalk 59: 192-218.
- Buhr, K.-J., Winter, J.E., (1976). Distribution and maintenance of a Lanice conchilega association in the weser estuary (FRG), with special reference to the suspension-feeding behaviour of Lanice conchilega in Biology of benthic organisms by Keegan, B.F., Ceidigh, P.O. and Boaden, P.J.S., Pergamon press: 101-113
- Camphuysen, C. J., van Franeker, J.A. (1992). The value of beached bird surveys in monitoring marine oil pollution. Report Vogelbescherming Nederland, Zeist.
- Camphuysen, C.J., Leopold, M.F. (1994). Atlas of seabirds in the southern North Sea. IBN Research Report 64/6. Institute for Forestry and Nature Research, Texel, 126 p.
- Cavalli-Sforza LL, Edwards AWF (1967). Phylogenetic analysis: models and estimation procedures. Am. J. Hum. Gen. 19: 233-257
- Carter, R.W.G. & Balsillie J.H. (1983). A note on the amount of wave energy transmitted over nearshore sand bars. Earth Surf. Proc. Land 8:213-222
- Cattrijsse, A., Dankwa, H.R., Mees, J. (1997). Nursery function of an estuarine tidal marsh for the brown shrimp Crangon crangon. J. Sea Res. 38: 109-121.
- Cattrijsse, A., Vincx, M. (2001). Biodiversity of the benthos and the avifauna of the Belgian Coastal waters. Summary of data collected between 1970 and 1998. The Prime Ministers's Services Federal Office for Scientific, Technical and Cultural Affairs. Brussel, 40 pp.
- Chow, S (1998). "Universal PCR primer for Calmodulin Gene Intron in Fish", Fish. Sci 64: 999-1000
- Christensen, H. & Kanneworff, E. (1985). Sedimenting phytoplankton as major food source for suspension and deposit feeders in the Oresund. Ophelia 24: 233-244
- Claeys D. (1979). Studie van het meiobenthos van de Kwinte Bank (Noordzee). Licentiaatsverhandeling RUG, 148 pp.
- Daan, N., Bromley, P.J., Hislop, J.R.G., Nielsen, N.A. (1990). Ecology of North Sea fish. Neth. J. Sea Res. 26: 343-386.
- Debacker, V., Holsbeek, L., Tapia, G., Gober, S., Joiris, C., Jauniaux, T., Coignoul, F., Bouquegneau, J.M. (1997). Ecotoxicological and pathological study of common guillemots Uria aalge, beached on the Belgian coast during six subsequent wintering periods (1989-90 to 1994-95). Diseases of aquatic organisms 29: 159-168.
- Degraer, S. (1999). Macrobenthos of shallow marine habitats (Belgian coast) and its use in coastal zone management. PhD thesis University Gent, Gent, 158pp.
- Degraer, S., Van Lancker, V., Moerkerke, G., Van Hoey, G., Vincx, M., Jacobs, P. & Henriet, J.-P. (2001). Intensive evaluation of the evolution of a protected benthic habitat: HABITAT. Final report 30/09/01. Federal Office for Scientific, Technical and Cultural Affairs (OSTC).
- De Putter, G., Orbie, G. (1990). Het voorkomen van de Dwergstern Sterna albifrons aan de Vlaamse kust. Mergus 4: 14-22.
- Dewicke, A. (2001). Hyperbenthic communities of the North Sea. Gent University, Phd thesis, 219 pp.
- Drgas, A., Radziejewska, T., Warzocha, J. (1998) Biomass size spectra of near-shore shallow-water benthic communities in the Gulf of Gdansk (Southern Baltic Sea). Mar. Ecol. PSZNI 19(3): 209-228
- Duplisea, D. (2000) Benthic organism biomass size-spectra in the Baltic Sea in relation to the sediment environment. Limnol Oceanogr 45(3): 558-568
- Duplisea, D.E., Drgas, A. (1999) Sensitivity of a benthic, metazoan, biomass size spectrum to differences in sediment granulometry. Mar. Ecol. Prog. Ser. 177: 73-81
- Dyer, K.R., Huntley, D.A. (1999) The origin, classification and modelling of sand banks and ridges. Cont. Shelf Res. 19: 1285-1330.
- Exadactylos, A., Geffen, A.J., Thorpe, J.P. (1998) Population structure of the Dover sole, Solea solea L., in a background of high gene flow. J. Sea Res. 40: 117-129
- Exadactylos A., Geffen A.J., Thorpe J.P., (1999) "Growth and genetic variation in hatchery-reared larval and juvenile Dover sole, Solea solea (L.)", Aquaculture 176, p.209-226
- Exadactylos A., Thorpe J.P. (2001). Allozyme variation and genetic inter-relationships between seven flatfish species (Pleuronectiformes). Zool.J.Linn. Soc. 132: 487-499.
- Furness, R.W. (1993). Birds as monitors of pollutants. In: Birds as monitors for environmental change. R.W. Furness & J.J.D. Greenwood (Eds.). Chapman & Hall, London: 86-143.
- Furness, R.W. and Greenwood, J.J.D. 1993. Birds as monitors for environmental change. Chapman & Hall, London, 343 p.
- Geets A. (1998). Host-parasite interactions between sympatric Pomatoschistus species (Gobiidae, Teleostei) and their helminth parasites: ecological and phylogenetic aspects. Doctoraatsthesis, Katholieke Universiteit Leuven, België .
- Geets A, Malmberg G, Ollevier F. (1998). Gyrodactylus longidactylus n.sp., a monogenean from Pomatoschistus lozanoi (de Buen, 1923) from the North Sea. Syst. Parasitol. 41, 63-70.
- Gläser H-J. (1974). Eine neue artengruppe des subgenus Gyrodactylus (Paranephrotus) (Monogenea, Gyrodactylidae). Zool. Anz 192, 271-278.
- Gheskiere, T. (2000). Structurele diversiteit van nematoden-gemeenschappen van de Bligh Bank (Zuidelijke bocht van De Noordzee). Licenciaatsverhandeling RUG; 92 pp + app.
- González-Oreja, J.A., Saiz-Salinas, J.I. (1998) Loss of heterotrophic biomass structure in an extreme estuarine environment. Est. Coast Shelf Sci. 48: 391-399
- Grioche, A., Koubbi, P., Harlay, X. (1999) Spatial patterns of ichthyoplankton assemblages along the eastern English Channel French coast during spring 1995. Estuar. Coast. Shelf S. 49: 141-152.
- Harris, PD. (1993). Interactions between reproduction and population biology in Gyrodactylid monogeneans - a review. Bulletin Français de la Pęche et de la Pisciculture 328, 47-65.
- Harris, M.P. (1997). Guillemot Uria aalge. In: The EBCC Atlas of European breeding birds - their distribution and abundance. Hagemeijer, W.J.M. and Blair, M.J. (Eds.). Poyser, London: 372-373.
- Hartmann-Schröder, G. (1996). Annelida, Borstenwürmer, Polychaeta - Tierwelt Deutschlands, Teil 58. Hamburg: Veb Gustav Fischer Verlag Jena. 1- 648
- Hebert, P. D. N. & Beaton, M .J. (1989). Methodologies for allozyme analysis using cellulose acetate electrophoresis: A practical handbook. Helena Laboratories Inc.
- Hildén, O., Tasker, M.L. (1997). Razorbill Alca torda. In: The EBCC Atlas of European breeding birds - their distribution and abundance. Hagemeijer, W.J.M. and Blair, M.J. (Eds.). Poyser, London: 372-373.
- Holtmann, S.E., Groenewold, A., Schrader, K.H.M., Asjes, J., Craymeersch, J.A., Duineveld, G.C.A., van Bostelen, A.J., van der Meer, J. (1996). Atlas of the zoobenthos of the Dutch Continental Shelf. Ministry of Transport, Public works and Water management, North Sea Directorate, Rijswijk, 1-244
- Hostens, K. (2000). Spatial patterns and seasonality in the epibentic communities of the Westerschelde (Southern Bight of the North Sea). J. Mar. Biol. Assoc. U.K. 80: 27-36
- Huston, M.A. (1994). Biological Diversity: the co-existence of species on
- changing landscapes. Houston (eds). Cambridge University Press., 681 pp.
- Iyengar, A., Piyapattanakorn, S., Heipel, D.A., Stone, D.M., Howell, B.R., Child, A.R. and Maclean N, (2000) "A suite highly polymorphic microsatellite markers in turbot (Scophtalmus maximus L.) with potential across several flatfish species", Mol. Ecol. , p.368-371
- Jarvis, P.J. (1993). Environmental changes. In: Birds as monitors for environmental change. R.W. Furness & J.J.D. Greenwood (Eds.). Chapman & Hall, London: 42-85.
- Katz, L. A., Harrison, R. G. (1997). Balancing selection on electrophoretic variation of PGI in two species of field cricket: Gryllus veletis and G. pennsylvanicus. Genetics 147: 609-621.
- Keijl, G.O., Leopold, M.F. (1997). Massaal fouragerende Dwergmeeuwen Larus minutus voor de Hollandse kust in april 1996. Sula 11: 17-20.
- Knijn, R.J., Boon, T.W., Heessen, H.J.L., Hislop, J.R.G. (1993) Atlas of North Sea Fishes. ICES Coop. Res. Rep. 194, 268 pp.
- Kocher, T.D., Thomas, W.K., Meyer, A., Edwards, S.V., Pääbo, S., Villablanca, F.X., A.C. Wilson (1989). Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proc. Nat. Acad. Sci. USA 86: 6196-6200.
- Kotoulas, G., Bonhomme, F., Borsa; P. (1995). Genetic structure of the common sole Solea vulgaris at different geographic scales Mar. Biol. 122: .361-375
- Leaper, R., Raffaelli, D., Emes, C., Manly, B. (2001). Constraints on body-size distribution: an experimental test of the habitat architecture hypothesis. J Animal Ecol 70: 248-259
- Lemaire, C., Allegrucci, G., Naciri, M., Bahri-Sphar, L., Kara, H., Bonhomme, F. (2000). Do discrepancies between microsatellite and allozyme variation reveal differential selection between sea and lagoon in the sea bass (Dicentrarchus labrax)? Mol. Ecol. 9: 457-467.
- Leopold, M.F., Baptist, H.J.M., Wolf, P.A. and Offringa, H. 1995. De Zwarte Zeeëend Melanitta nigra in Nederland. Limosa 68: 49-64.
- Lloyd, C., Tasker, M.L. and Partridge, K. 1991. The Status of Seabirds in Britain and Ireland. NCC-Seabird Group. Poyser, London.
- Maes, F., Cliquet; A., Seys, J., Meire, P., Offringa, H. (2000). Limited Atlas of the Belgian part of the North Sea. OSTC, Brussels, 31 pp.
- Mantel, N. (1967). The detection of disease clustering and generalized regression approach. Canc. Res. 27: 209-220.
- McKay, S.I. and Miller, P.J. (1997). The Affinities of European Sand Gobies (Teleostei: Gobiidae). J. Nat. Hist. 31: 1457-1482.
- Miller, P. J. (1986) Gobiidae. In: Fishes of the north-eastern Atlantic and the Mediterranean. Volume III. p. 1018-1085.
- Montevecchi, W.A. (1993) Birds as indicators of change in marine prey stocks. In: Birds as monitors for environmental change. R.W. Furness & J.J.D. Greenwood (Eds.). Chapman & Hall, London: 217-266.
- Mork, J. & Sundnes, G. (1985). O-group cod (Gadus morhua) in captivity: differential survival of certain genotypes. Helgol. Meeresunt. 39: 63-70.
- Nei, M. (1978). Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89: 583-590.
- Offringa, H., Seys, J., Van den Bossche, W. , Meire, P. (1996). Seabirds on the Channel doormat. De giervalk 86: 3-71.
- Penzo, E., Gandolfi, G., Bargelloni, L., Colombo, L., Patarnello, T. (1998). Messinian Salinity Crisis and the Origin of Freshwater Lifestyle in Western Mediterranean Gobies. Molecular Biology and Evolution. 15: 1472-1480.
- Peters RH (1983) The ecological implications of body size. Cambridge University Press, Cambridge
- Powers, D. A., Lauermann T., Crawford D., DiMichele L. (1991). Genetic mechanisms for adapting to a changing environment. Annual Review of Genetics 25: 629-659.
- Raymond, M. & Rousset, F. (1995). GENEPOP version 3.1: population genetics software for exact tests and ecumenism: updated version of Genepop 1.2. J. er. 86: 248-249.
- Rice, W. R. (1989). Analyzing table of statistical tests. Evolution 43: 223-225.
- Richardson, P. J., Baverstock, P. R., Adams, M. (1986). Allozyme electrophoresis: A handbook for animal systematics and population studies. Academic Press Inc., London.
- Ridley, M. (1996). Evolution. Blackwell Science, Cambridge
- Rohde, K. (1996). Robust phylogenies and adaptive radiations: a critical examination of methods used to identify key innovations. American Naturalist 148: 481-500
- Rousseau, V. (1999) Dynamics of Phaeocystis and diatom blooms in the eutrophicated coastal waters of the Southern Bight of the North Sea. Phd thesis. pp. 204
- Sajdak, S.L. and Phillips, R.B. (1997) Phylogenetic relationships among Coregonus species inferred from the DNA sequence of the first internal transcribed spacer (ITS1) of ribosomal DNA. Can. J. Fish. Aquat. Syst. 54: 1494-1503
- Shanks, A.L. (1995) Mechanisms of cross-shelf dispersal of larval invertebrates and fish. In: McEdward, L. (ed.). Ecology of marine invertebrate larvae, CRC Press, Boca Raton, pp. 323-367.
- Seys, J., Meire, P., Kuijken, E. (2001a). Focal species and the designation and management of marine protected areas: sea- and coastal birds in Belgian marine waters. In: Sea- and coastal bird data as tools in the policy and management of Belgian marine waters. Seys, J. (Ed.). Proefschrift Universiteit Gent, Gent: 40-67.
- Seys, J., Vantieghem, S., Offringa, H., Meire, P,. Kuijken, E. (2001b). Monitoring seabirds on fixed ferry routes: a methodological evaluation. In: Sea- and coastal bird data as tools in the policy and management of Belgian marine waters. Seys, J. (Ed.). Proefschrift Universiteit Gent, Gent: 68-77.
- Seys, J., Offringa, H., Van Waeyenberge, J., Meire, P. and Kuijken, E. 2001c. Long-term changes in oil pollution off the Belgian coast: evidence from beached bird monitoring. In: Sea- and coastal bird data as tools in the policy and management of Belgian marine waters. Seys, J. (Ed.). Proefschrift Universiteit Gent, Gent: 97-108.
- Seys, J., Offringa, H., Van Waeyenberge, J., Meire, P., Kuijken, E. (2001d). An evaluation of beached bird monitoring approaches. In: Sea- and coastal bird data as tools in the policy and management of Belgian marine waters. Seys, J. (Ed.). Proefschrift Universiteit Gent, Gent: 109-122.
- Seys, J., Offringa, H., Van Waeyenberge, J., Meire, P., Vincx, M., Kuijken, E. (2001e). Distribution patterns of seabirds in Belgian marine waters. In: Sea- and coastal bird data as tools in the policy and management of Belgian marine waters. Seys, J. (Ed.). Proefschrift Universiteit Gent, Gent: 23-39.
- Schneider, S., Roessli D. & Excoffier L. (2000). ARLEQUIN ver 2.000: A software for population genetic analysis. Genetics and Biometry Laboratory, University of Geneva, Geneva
- Shaklee, J. B., Allendorf, F. W., Morizot D. C. & Whitt G. S. (1990). Gene nomenclature for protein-coding loci in fish. Trans. Am. Fish. Soc 119: 2-15.
- Sotelo, C.G., Calo-Mata, P., Chapela, M.J., Perez-Martin, R.I., Rehbein, H., Hold, G.L., Russell, V.J., Pryde, S., Quinteiro, J., Izquierdo, M., Rey-Mendez, M., Rosa, C., Santos, A.T. (2001) Identification of flatfish (Pleuronectiforme) species using DNA-based techniques. J. Agric. Food Chem. 49: 49: 4562-4569
- Steyaert, M., Garner, N., Van Gansbeke, D., Vincx, M. (1999). Nematode communities from the North Sea: environmental controls on species diversity and vertical distribution within the sediment. J. Mar. Biol. Ass. U.K. 79:253-264
- Swofford, D. L. (1999). PAUP*: Phylogenetic Analysis Using Parsimony (and other methods), version 4.0. Massachussetts: Sinauer Associates.
- Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting positions-specific gap penalties and weight matrix choice. Nuc. Acid Res. 22: 4673-4680.
- Trentesaux, A., Stolk, A., Berné, S. (1999) Sedimentology and stratigraphy of a tidal sand bank in the southern North Sea. Mar. Geol. 159: 253-272.
- Turan, C., Carvalho, G. C. & Mork, J. (1998). Molecular genetic analysis of Atlanto-Scandian herring (Clupea harengus) populations using allozymes and mitochondrial DNA markers. J. Mar. Biol. Ass. U.K. 78: 1-15.
- Vanaverbeke, J., Gheskiere, T., Vincx, M. (2000). The meiobenthos of subtidal sandbanks on the Belgian Continental Shelf (Southern Bight of the North Sea). Est. Coast. Shelf Sci. 51: 637-649
- Vanaverbeke, J., Steyaert, M., Bonne, W., Vincx, M. (2001). Three decades of meiobenthos research on the Belgian continental shelf: an overview. In: Cattrijsse A. & Vincx M. (eds.) Biodiversity of the benthos and the avifauna of the Belgian coastal waters. The Prime Ministers's Services Federal Office for Scientific, Technical and Cultural Affairs. Brussel.11-20
- Vanaverbeke. J., Steyaert, M., Vanreusel, A., Vincx, M. (subm) Nematode biomass spectra as a descriptor of functional changes in nematode communities due to human and natural induced impact.
- Van Hoey, G., (2000). Benthisch-pelagische koppeling: structurele en functionele respons van het macrobenthos van twee stations in de Noordzee. Licentiaatsverhandeling RUG. 125 pp
- Van Hoey, G., Degraer, S., Vincx, M., (in prep). The macrobenthic communities of the Belgian Continental Shelf: a (re)analysis of combined data over six year.
- Van Impe, J. (1966). Note sur la migration d'automne de Larus minutus sur la côte belge. Alauda 34: 63-65.
- Venkatesh B., Ning Y. and Brenner S. (1999) Late changes in spliceosomal introns define clades in vertebrate evolution. Proc. Natl. Acad. Sci. USA96, p.10267-10271
- Volckaert, F.A.M., Kayens, G., Schallier, R., Jacques, T.G. (2000). Aerial surveillance of operational oil pollution in Belgium's maritime zone of interest. Mar. Poll. Bull. 40: 1051-1056.
- Wallis, G. P. & Beardmore, J. A. (1984a). An electrophoretic study of the systematic relationships of some closely related goby species (Pisces, Gobiidae). Biol. J. Linn. Soc. 22: 107-123.
- Wallis, G. P. & Beardmore, J. A. (1984b). Genetic variation and environmental heterogeneity in some closely related goby species. Genetica 62: 223-237.
- Walsh, P.S., Metzger, D.A., Higuchi, R. (1991) Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques: 10, 506-513.
- Watt, W. B. (1983). adaptation at specific loci. II. Demographic and biochemical elements in the maintenace of the Colias PGI polymorphism. Genetics 103: 691-724.
- Weir, B. S. (1990). Genetic data analysis. Sinauer Sunderland, MA.
- Weir, B. S. & Cockerham, C. C. (1984). Estimating F-statistics for the analysis of population structure. Evolution 38: 1358-1370.
- Willems K., Vincx M., Claeys D., Vanosmael C. & Heip C. (1982). Meiobenthos of a sublittoral sandbank in the Southern Bight of the North Sea. J. Mar. Biol. Ass. U.K., 62: 535-548.
- Woehrling, D. (1985) Données sur la population de soles du secteur de Gravelines (Nord). Contribution ŕ la connaissance des oeufs, larves et juvéniles. Rapport IFREMER pour EDF, 2 vol., 237 pp.
- Woehrling, D., Le Fčvre-Lehoërff, G. (1998) Long-term series in ichthyoplankton: Sole and sprat at French coast of the North Sea from 1975 to 1994. Oceanol. Acta 21(1): 113-121.
- Wright, S. (1951). The genetic structure of populations. Ann. Eugen. 15: 323-354.
- Zietara, M.S. et al. (2000). The nuclear rDNA region of Gyrodactylus arcuatus and G. branchicus (Monogenea: Gyrodactylidae). J. Parasitol. 86(6): 1368-1373.
|
General coordination: Magda Vincx, Jan Vanaverbeke Web site and databases hosted by VLIZ |
